Human Perception of Visual Information (3)
Chap3?Affective Perception: The Power Isin the Picture
1 Introduction
自从照相机发明以来,摄影师就利用图片的力量来引发观众的情感反应。摄影记者捕捉战争和暴力的场景,令人兴奋和恐惧;营销机构寻求能促使消费者出席并购买的形象;公共卫生运动利用照片瞄准和鼓励更安全的行为;慈善组织邮寄图片请求救助的;各种各样的媒体呈现的场景旨在刺激和唤起。从基础科学的观点来看,照片图像(因为它们在知觉上与所描绘的事件非常匹配)作为显著的检索线索,激活了与调节情绪体验的皮层下和皮层区域的关联联系的现有心理表征。因此,情绪场景感知已被证明是一种有效的方法,用于情绪的基础研究,以及各种神经、精神和其他心理健康障碍中的情绪功能障碍(例如,de Tommaso等人,2009;De Zorzi等2021;Hermanns等人2003;Huijbers等2011;Ihssen等2011;Renfroe et al . 2016)。
在本章中,我们简要回顾了我们之前对情绪场景感知的研究,并利用现有数据库进行了新的基于图片的分析,以评估个体场景可靠地参与个体情绪反应的程度。我们首先简要地考虑情绪的动机观点(1.2),然后描述现有的数据库(1.3)和分析策略(1.4),以评估图片的可靠性。然后计算情绪体验(1.5.1)、皮肤电导(1.5.2)、瞳孔直径评估报告的场景可靠性(1.5.3)、面肌电图(1.5.4)、心率(1.5.5)、杏仁核功能活动(1.5.6)和自由回忆记忆(1.5.7)。我们得出结论(1.6),在测量中,特定的图像可靠地引发情绪反应,并讨论了情绪反应的异同,因为它们随享乐内容和依赖测量而变化。
2 Motivation and Emotion
英语中有数百个单词描述了各种各样的人类情感体验(例如,愤怒、恐惧、快乐、悲伤等),几十年来,科学家们提出了不同的标签列表,这些标签被认为是情感研究的基础(例如,Ekman 1971;Plutchik 1980)。基于动物研究和理论(例如,Dickinson & Dearing 1979;Konorski 1967),我们从一个更简单的方法开始,其中两种动机系统被认为是情感体验的基础。这两个系统——防御系统和食欲系统——在神经回路中执行,这些神经回路在进化上是古老的,在哺乳动物物种中是共享的,并且已经进化到调节各种维持和保护生命的行为。防御系统主要在生命受到威胁的情况下被激活,具有建立在撤退,逃跑和攻击上的原始行为曲目;食欲系统在促进生存的环境中被激活,包括维持、繁殖和养育,并具有摄食、交配和照顾等基本行为。
根据这种观点,情绪体验的基本特征是防御或食欲激活的强度,我们认为这是情绪体验的战略方面(Lang & Bradley 2010;Lang et al . 1997;L?w et al . 2008),通常有相似的目标,即引导注意力和准备行动,而不管动机背景如何(Bradley 2009)。而且,尽管基本的接近和退缩的双向行为代表了简单物种的行为库(例如,应用,Schneirla 1959),但在更复杂的生物中,已经进化出了大量的应对行为库,以保护生命并允许其繁殖。这种情绪的战术方面可以在不同的动机背景下促使类似的行动,也可以在相同的动机背景下促使不同的行动(Lang & Bradley 2013)。
从外行人的角度来看,情绪主要涉及内部感觉状态,这是无法通过科学测量的。相反,正如Lang (1968, 2010;Lang & Bradley 2018,参见Hugdahl 1981)最初提出的,情绪体验可以用三个系统来衡量:(1)主观报告/表达语言,(2)生物和生理反应,(3)公开行动。没有一个单一的反应系统与假设的意识感觉状态有一对一的关系——一个人可以在没有其他可测量的防御激活指数的情况下报告高度恐惧,而另一个人在可测量的防御激活的情况下报告没有痛苦。而不是理解有意识的情绪体验,情绪的实证研究只是旨在表征和描述三系统反应,因为它们在特定的情绪诱导环境中是不同的(Lang 2010;布拉德利2000)。
3 Emotional Reactivity Database
在过去的许多年里,我们的情绪研究的一个目标是描述场景感知背景下的情绪反应。从实验的角度来看,摄影图像的静态特性是理想的,因为情绪反应的许多生物和生理指标(如心率、皮肤电导等)对动态刺激变化(如剪切、平移、变焦等)很敏感,这可能会干扰检测特定于情感内容的反应(参见Detenber & Lang 2010的概述)。首先,我们从各种可用的来源收集了一组情感唤起和中性的照片,并从一大群参与者那里收集了快乐和唤醒的报告(国际情感图片系统,IAPS: Lang et al 2008)。快乐和觉醒的判断被认为是动机激活的两个参数,有关于动机系统参与的快乐指数(即防御或食欲)的报告,以及对其强度的觉醒判断。
初步研究(见Bradley & Lang 2007;Lang(2010)在自由观看情境下呈现了情绪和中性的场景,发现愉快和不愉快的场景会引起皮肤电导和瞳孔直径变化的增强,杏仁核(食欲和防御动机系统的关键皮质下节点)的激活增强,以及更好的自由回忆。另一方面,在愉快和不愉快的场景中,心率变化、惊吓反射和面部肌电图活动有所不同。然而,后来的研究发现,尽管对快乐和/或兴奋的评级相似,但特定的场景内容在情绪反应的大小中起着重要作用,这表明评估评级虽然高度可靠,但并不是情绪投入的直接解读(见布拉德利等人)2001年,2017年)。
本章的分析使用了我们实验室先前发表和未发表的许多研究的数据,包括每个参与者的愉悦和唤醒的评估报告,变化分数(从图像开始前的基线开始)和皮肤电导,心率,面部波纹肌和颧肌肌电活动的场景(72张图片;Bradley等人
2001a),瞳孔直径(60张;Bradley等人2017b),杏仁核的功能活动(50张图片;未发表的数据)和自由召回(360图片;Bradley等人2017年)。
在所有的研究中,每张图片都是新颖的,每个参与者只看一次;每项研究还包括一组中性场景。特定场景呈现的顺序在参与者之间得到了平衡,减少了仅仅由于输入序列位置而导致的情绪反应差异。在研究中,我们集中于描绘色情、浪漫、冒险、运动、家庭、食物、自然、污染(如厌恶)、威胁和残害场景的具体内容(见图1)。

4 Analytic Plan
当前分析的核心兴趣是,对于各种不同的3系统情绪测量,特定场景如何可靠地诱发参与者的情绪反应。之前的分析平均了每个参与者对图片的情绪反应,而在当前的分析中,我们平均了参与者对每张图片的情绪反应。一种可能性是,一般来说,参与者对特定内容的不同范例(例如,威胁)有特殊的反应,在个体之间诱导的情绪反应中几乎没有共性,这预示着整体场景可靠性较低。另一方面,特定场景可能会引起个体情绪反应的增强(或减弱),从而导致高场景可靠性。此外,对于情绪反应的某些测量,作为特定场景函数的可靠性可能高于其他测量。
单次试验(图片)分析需要一个无测量的情绪反应指标,该指标将每个场景、每个参与者的情绪参与程度指数化,而不考虑所测量的情绪反应。为了做到这一点,在每次试验中,参与者的反应(每个测量单独)都是基于个体对一组中性场景的反应的平均值和标准偏差进行标准化的。这些情绪反应(z)分数(以标准偏差的自由度量单位表示),对每个人的反应程度进行指数,比中性场景引起的反应更大(正z分数)或更少(负z分数),并在个体之间进行平均,以产生每个场景的情绪反应分数。
为了评估图像可靠性,对每个依赖测量进行了排列分析,其中参与者被随机分为两个亚组,并计算每个亚组中每张图片的平均情绪反应性(z)得分。
然后,计算Pearson积差相关性(r),索引相同的图片在队列中与或多或少的情绪反应相关的程度。对于每个测量,排列分析进行了500次,这些相关性的结果平均值和分布被用来估计特定场景在每个情绪反应测量中可靠地引起个体情绪反应的程度。平均情绪反应(z)得分也在每个图片内容的样本中平均,并在单变量方差分析中评估特定场景内容的总体效果,随后通过LSD比较检验来评估场景内容之间的差异。
在第二个可靠性分析中,通过计算每个测量中出现在情绪反应(z)分布上半部分的场景的比例,来评估每个内容类别中促使强烈情感投入的示例数量。如果场景内容是情感投入中需要考虑的重要因素,那么持续引发情感反应的场景所占比例应该更高。
5 Affective Scene Perception
5.1 Evaluative Reports
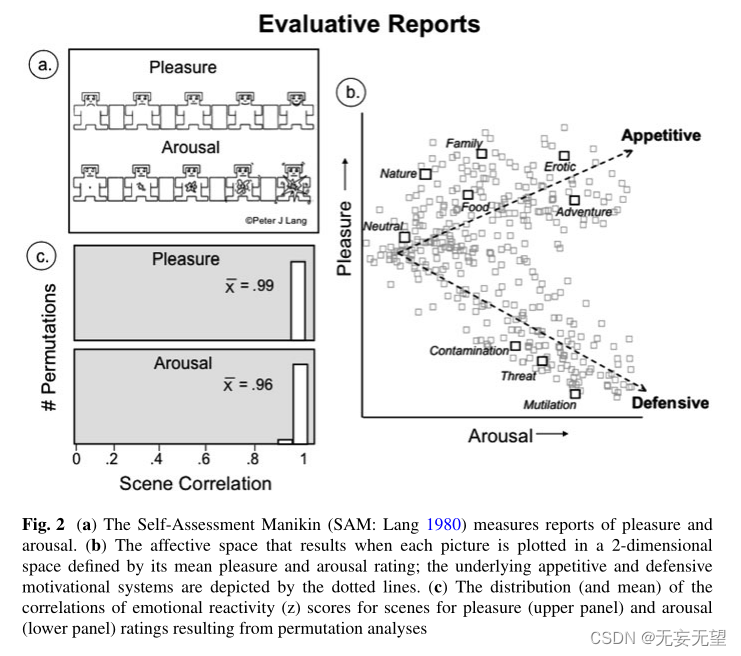
为了测量快乐和觉醒的报告,Lang(1980)开发了自我评估模型(SAM;见图2a)一个图形评定量表,它提供了一种无语言、无文化、无年龄的评估报告测量方法,与管理更长的语义差异产生相同的结果
图2b说明了根据唤起的愉悦感和兴奋感对图片进行评级时产生的情感空间:被评为高度愉悦或高度不愉悦的场景与较高的兴奋感评级相关,这促使愉悦感和兴奋感之间形成了显著的二次关系。图2c中的虚线说明了被评估的愉悦感和兴奋感与假设的潜在的食欲和防御动机系统的关系,分别有不同激活强度的食欲和防御动机系统。
图2c显示了情绪反应性(z)评分的排列分析结果。关于愉悦感和兴奋感的评估报告在整个队列中都是高度可靠的,愉悦感的平均场景相关性为0.99,兴奋感的平均场景相关性为0.96,两者都是非常显著的。因此,尽管存在一些微小的差异,但在整个生命周期(Ferrari等人2017年)和文化(例如Molto等人1999年)中,男性和女性的情感空间形状是一致的(Bradley等人2001年)。Lasaitis等人2008;V erschuere et al . 2001),并且与其他情感线索的情感空间形状相同,包括声音、单词和文本(Bradley & Lang 2007)。
5.2 Skin Conductance?
皮电活动是最早用来衡量情绪反应的心理生理指标之一。尽管多种生理反应是由副交感神经系统和交感神经系统活动双重支配的,但皮电活动仅由交感神经系统支配,其激活介导了众所周知的“战斗或逃跑”反应。一个皮肤电指数——皮肤电导活动——是通过在两个传感器之间发送一个不可检测的电流来测量的(通常放在手上,见图3a)。交感神经系统的激活降低了皮肤对电流的抵抗力,提高了皮肤的电导活性。在早期的图片感知研究中,当人们看到被评为情绪化的图片时,皮肤电导的增加是可靠的,而不是中性的,无论场景在享乐价中被评为愉快还是不愉快(例如Greenwald等人1989;Winton et al . 1984)。交感神经系统并不是简单地对厌恶的刺激作出反应,它也可靠地参与强烈的食欲激活,推测它不仅为个体做好了战斗和逃跑的准备,而且也为适当的食欲行为做好了准备(Bradley 2009)。
皮肤电导的排列分析(见图3b)表明,特定场景引发皮电活动的可靠性相当高,随机队列的平均场景相关性为0.71 (p < 0.05)。当对特定类别的样本进行平均时(见图3c),所有情绪内容引起的皮肤电导反应明显大于中性场景(即z = 0;单尾p < 0.05),色情、威胁和残害场景与其他场景内容相比,显著提高了反应。
此外,作为如图3d所示,虽然没有家庭、食物或自然的例子是最被动的场景,但超过80%的色情、切割和威胁例子在分布的上半部分引发情绪反应得分。虽然在大多数反应性场景,这些示例的平均情绪反应性(z)得分(见图3d中条形图上方的数字)与整体情绪较低一致这些场景内容引发的反应

虽然特定的场景确实会引起个体的皮肤电导活动,但这种测量方法可能不是情绪反应的最佳指标。人们早就知道,相当大比例的参与者(约40%)没有反应——在整个实验中,皮肤电活动没有可测量的变化(V使能和米切尔,1996年)。此外,皮肤电导对新颖性高度敏感(Bradley 2009),在整个实验过程中迅速适应,导致许多零试验。另一种测量交感神经系统激活的方法——瞳孔直径——更有希望。?
5.3 Pupil Diameter
瞳孔直径的变化受双肌控制——括约肌和扩张肌——它们分别受副交感神经和交感神经系统活动的支配。Hess和Polt(1960)提出,瞳孔在积极的场景下会扩大(“打开”),在消极的场景下会缩小(“关闭”),这与瞳孔反应的接近或回避的假设是一致的,但这些早期研究中的方法和测量问题导致无法复制。我们的早期研究发现,在观看图片时,瞳孔扩张对食欲和防御激活的强度都很敏感,在愉快和不愉快的场景下瞳孔扩张的增强与皮肤电导反应性高度相关(Bradley et al . 2008;Henderson et al . 2014)。
在场景感知期间测量的瞳孔波形通常包括最初的收缩,随后是随后的扩张。该波形的因子分析确定了两个独立的因素,包括由场景亮度调节的初始收缩(可能由副交感神经系统激活介导)和由交感神经介导的场景情绪调节的后期扩张(Bradley et al . 2017b)。由于最初的光反射主要对场景亮度和亮度的差异做出反应,因此在观看图像时仔细控制这些变量至关重要。可能的控制包括通过亮度匹配每个内容的特定场景;等于所有场景的亮度,和/或在场景观看之前控制瞳孔的直径(例如,匹配即将到来的场景亮度)。
然而,尽管有这些控制,我们发现瞳孔直径也会受到注视时局部区域亮度的影响。例如,一个有云的场景,在整个观看过程中,瞳孔直径会变小,因为前景信息(云)比背景(天空)更亮,尽管整个场景的亮度与其他刺激是相同的。一种解决方案是通过减小图像尺寸来呈现每个场景,以便在每次固定时都能看到整个图像(例如,所有亮度级别),尽管这种操作可能会影响对情感内容的理解(例如,De Cesarei & Codispoti 2010)。相反,我们在5×5矩阵中呈现场景(等同于整体亮度),其中相同的场景在所有25个单元中重复(见图4a)。场景内容不仅易于感知,而且这种操作在消除因注视时图像亮度差异而导致的瞳孔直径差异方面非常有效(Bradley等,2017b)。
图4b显示,晚期瞳孔扩张的可靠性相当高,随机队列的平均场景相关性为0.71 (p < 0.05)。正如皮肤电导所发现的那样,与观看中性场景相比,观看任何情绪内容都会引起更大的瞳孔扩张(见图4c)。观看色情场景时瞳孔放大的程度与观看威胁和残害场景时瞳孔放大的程度相同,观看污染场景时瞳孔放大的程度也相似。与所有其他场景内容相比,色情和残害场景引起了更大的扩张,这些内容中最具反应性的范例比例很高(见图4d)。

总的来说,与皮肤电导相比,在后期瞳孔扩张分布的上半部分,跨情感类别的样本比例更大,这表明这种测量可能对交感参与的更温和的差异敏感。尽管与皮肤电导不同,相当大比例的污染样本是最具反应性的场景(见图4d),但两种测量方法的具体样本(和参与者)不同。未来的研究,包括测量和相同的例子在一个主题内的设计可以解决这个问题。
5.4 Facial EMG
面部对情绪事件的反应,如皱眉和微笑,是明显的行为主要在现实世界中扮演社会沟通的角色,但很微妙面部肌肉活动的变化可以通过使用放置在适当面部肌肉上的电极监测肌电图(EMG)活动来检测单独观看。
最常见的是,测量活动的方法是将传感器放置在皱眉时活跃的波纹肌和微笑时活跃的颧大肌上。最初的研究发现,与中性图片相比,在观看不愉快的图片时,皱襞肌的活动显著增加(Cacioppo et al . 1986),与交感神经调节的皮肤电导和瞳孔直径变化不同,皱襞肌肌电图活动增强是在不愉快的场景中发现的,而在愉快的场景中,皱襞肌的松弛程度通常低于基线(Lang et al . 1993)。
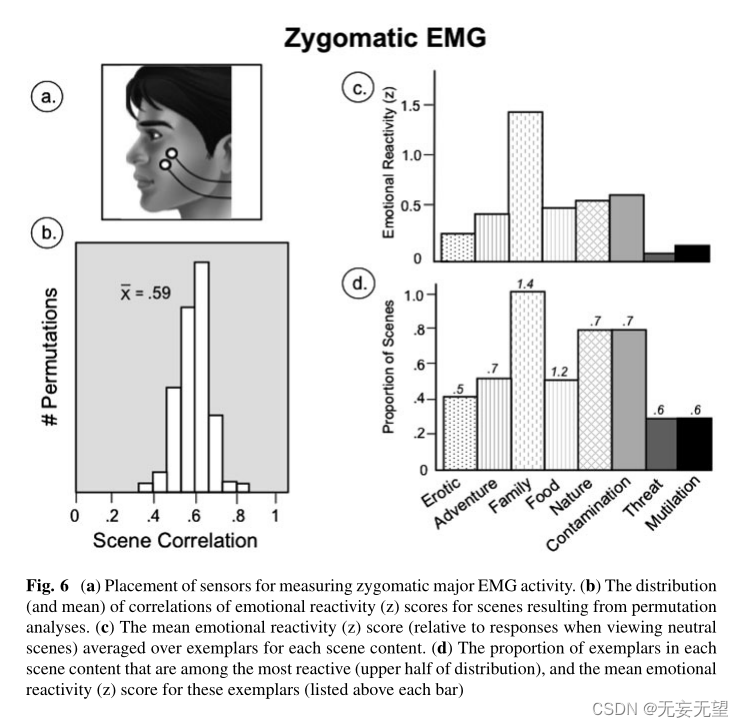
图5a显示了测量面部瓦楞肌肌电图活动时电极的位置。波纹肌肌图的排列分析(见图5b)表明,特定场景可靠地引发波纹肌肌图反应,随机队列的总体平均场景相关性为0.76 > 0.71 (p < 0.05)。对于所有令人不快的场景内容,皱襞肌肌电活动显著增强,而对于冒险、食物和家庭场景,皱襞肌肌电活动显著减少。切割场景比所有其他内容都更能刺激面部肌肉活动,威胁和污染场景也更能刺激
明显大于除情色场景以外的所有内容的波纹器活动。
当评估特定的例子时(图5d),所有令人不快的内容的大部分场景出现在那些引起最高波纹肌电反应的场景中。尽管在最具反应性的场景中出现了相对较大比例的情色场景,但这些例子的平均情绪反应(z)得分(即0.3)明显低于不愉快的场景。

图6a说明了电极放置在颧大肌上,当脸颊收缩或收紧时,如微笑时,颧大肌被激活(Tassinary et al 1989)。引起颧肌电图活动的特定图像的可靠性是显著的,随机队列的平均场景相关性为0.59 (p < 0.05;图6 b)。当对样本进行平均(见图6c)时,与观看中性场景相比,色情、家庭、冒险和自然的愉快场景显著提高了颧骨肌电活动的变化(p < 0.05),只有家庭场景比其他愉快内容引起显着更高的情绪反应(z)得分。虽然平均颧骨(z)对于令人愉悦的食物图片,反应性在数值上很大,但没有达到显著性(p = 0.15),可能是因为这种效应在女性中最为明显(Bradley et al . 2001)。在颧骨肌电信号活动中最活跃的具体例子(图6d)包括所有的家庭场景,其中描绘婴儿和儿童,经常微笑,这可能暗示面部模仿在部分或全部颧骨肌电信号变化中起作用。
另一方面,相当大比例的不愉快的污染场景(约70%)也会引起颧肌肌电活动的显著变化(高于中性),如果面部肌肉的活动仅仅是微笑的指标,这是出乎意料的。
这些数据说明了面部肌电图活动的两个要点。首先,面部肌肉活动并不局限于单一的表情。例如,表达厌恶的鬼脸可能涉及到瓦楞肌和颧肌的活动,从而导致在某些情感情境下的共同激活(Bradley等人,2001)。
除了污染场景,情色场景也会引起波纹肌和颧肌肌电图的共同激活,其中一些例子在两种测量中都是最活跃的。一种假设可能是,对于某些主题(或场景),色情图片是防御性的,而不是性欲的,吸引人的。另一方面,面部肌电图反应是少数由个人自愿控制的身体反应之一,可以反映文化或政治规范,这些规范虽然本质上是交际性的,但不一定反映基本的动机激活。
?5.5 Heart Rate
情绪图像感知的早期研究(如Libby et al . 1973;K·l·m·a·m·a·d·1975;Greenwald等人(1989)一致发现,当人们看到不愉快的场景时,心率会减慢(即减速)。基于这些数据,Lacey(1967)假设心脏减速指数增加了对厌恶线索的感觉摄入,并且,当后来评估特定图像内容的影响时,心脏减速不仅对威胁和残害的高度唤起的图像有效,而且对所有令人不快的内容都有效。当我们评估特定图片内容的影响时,不仅对威胁和残害的高度唤起的图片,而且对污染和损失的令人不快的场景也有增强的心脏减速,这些场景通常被认为是唤醒较低的(Bradley等)
2001年;Gomez & Danuser 2010)。在厌恶的图片观看过程中,心脏减速让人联想到在动物对威胁线索做出反应时发现的“恐惧性心动过缓”(例如,Campbell等人,1997年),这是迷走神经介导的,并被解释为反映了一种“停下来、看一看、听一听”的姿态,增强了感觉摄入和定向(Lacey 1967;格雷厄姆1979)。与感官摄入增加的假设相一致,后来的研究发现,高度兴奋的愉快图片,特别是那些涉及色情的图片,也会引起显著的初始心脏减速
(2001a)并且当重复呈现相同的图像时,所有的减速心脏反应都大大减少或不存在,从而减弱了知觉加工要求(Bradley等,1993;布拉德利2009)。
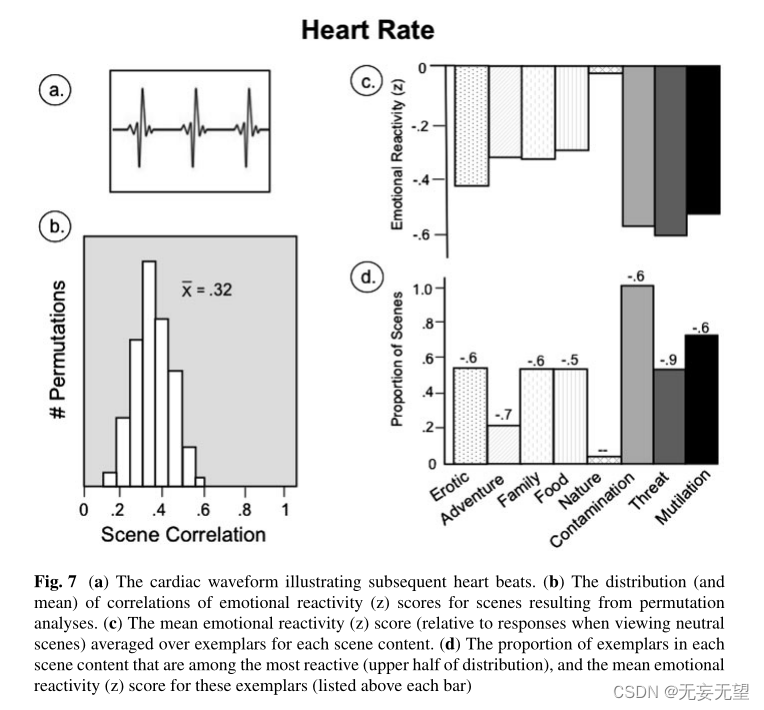
心率通常用心电图测量,产生如图7a所示的众所周知的心脏波形。后续节拍之间的时间间隔长度通常脱机转换为每分钟的节拍数。与其他一些生理指标不同,心脏通常每时每刻都在忙于与保持身体活力相关的许多活动,并且与呼吸和姿势等其他生理系统的变化密切相关,使其对情绪或认知因素的敏感性更加多变。因此,如图7b所示,场景中心脏变化的可靠性略低于其他生理测量,随机队列中的平均场景相关性为0.32,但仍然具有统计学意义(p < 0.05)。
图7c显示了在每个内容类别的样本中平均的心率变化的情绪反应(z)分数。除自然和家庭场景外,所有场景内容都有显著的心脏减速(相对于中性场景观看)(单尾p = 0.06)。心脏反应性(z)评分用于污染,威胁和残害场景,并且比自然场景(以及所有其他内容)更大。尽管色情场景与不愉快的内容一样会导致减速,但不愉快的威胁和污染场景比家庭、食物或冒险场景更能导致减速。与早期的数据一致,表明不愉快的场景会导致心脏大幅减速,每个不愉快类别中相当大比例的样本都是最具反应性的图片(见图7d)。
没有一个自然的例子在心脏减速方面具有强烈的唤起性(与整体内容缺乏反应性一致),而情色,家庭和食物的特定例子是最具反应性的,其情绪反应性(z)得分与不愉快的场景相当。综上所述,心脏减速,可能是索引定向和感觉处理,在情感场景感知中是重要的,并且在不愉快的场景,以及色情场景,以及其他一些令人愉快的内容的例子中得到加强。
5.6 Functional Amygdala Activity
啮齿类动物模型表明,杏仁核在两种防御机制中都起着调节作用(例如,Davis 1989;勒杜1995;Davis & Shi 2000)和食欲行为(Ishikawa et al 2008),以及对人类和非人类灵长类动物的研究支持杏仁核在情感视觉感知过程中通过一系列与视觉皮层的投射而发挥核心作用(Amaral et al 1992;Finke等人2019;Sabatinelli et al 2009, 2005;Vuilleumier et al . 2004)。双侧杏仁核在皮层下深处构成小的杏仁状区域(图8a),与海马(记忆)、额顶叶(注意)、岛(自主神经加工)和运动区域双向连接,构成支持防御或食欲作用的关键枢纽。最近的研究一致发现,与中性场景相比,当参与者看到愉快或不愉快的场景时,双侧杏仁核的功能激活会增强(例如Chang et al . 2015;Sabatinelli et al . 2009;Sambuco等2020;Wilson et al . 2020)。
个别场景对杏仁核激活的可靠性不如某些测量方法高,但仍然显著,平均场景相关系数为0.46 (p < 0.05);图8 b)。从场景内容的平均来看(见图8c),与中性场景观看相比,描绘色情、浪漫、威胁和残害的图片会显著增强杏仁核活动的功能(见图8c),色情和残害的场景表现出较高的反应性,两者没有区别。与反应最小的内容(冒险)相比,浪漫和威胁的场景也显示出增强的杏仁核激活。

另一方面,杏仁核在污染场景下的激活,会引起整体的高反应性,与在中性场景下的激活没有显著差异,这表明在不同的例子中存在显著的差异。
与这一假设相一致,图8d表明,大约一半的污染样本确实出现在分布的上半部分(这些特定场景的平均情绪反应性(z)得分相对较高)。另一方面,与杏仁核激活最多相关的具体例子,在很大程度上,是促使杏仁核激活最高的内容的成员,包括色情,威胁和残害场景。也许有一个例外,那就是相对较高比例的浪漫场景是最具反应性的场景。
5.7 Free Recall
与观看场景时唤起的三系统情绪反应不同,对情绪场景的记忆是初始认知和情感加工的后续结果。动物研究已经证明,学习和记忆受益于,并且潜在地依赖于,显著的动机激活,因为如果不是不可能的话,在缺乏动机命令(奖励或威胁)的情况下研究动物的认知是很困难的,因为动机命令被用来引导注意力和记忆。多年来,人类对情感唤起事件的记忆引发了多种假设,包括“积极”偏见,认为记忆偏向于愉快的事件(例如,“玫瑰色的眼镜”;Matlin & Stang 1978;Thompson 1985),以及一种“消极偏见”,这种偏见认为不愉快的事件只会被很好地记住(例如,“闪光灯记忆”;Brown & Kulik 1977;Bohannon 1988;Christianson & lotus 1987;Harris & Pashler 2005)。然而,数据和理论都支持一个假设,即情绪唤起事件,无论是愉快的还是不愉快的,都与增强的记忆表现有关(例如,Craik & Blankstein 1975;艾森克1976;W . a . k . e.(1958)),并且,与这些早期数据一致,许多评估记忆性能的研究报告称,愉快和不愉快的场景都能带来更好的记忆(例如,Bradley等人,1992;哈曼2001;McGaugh 2004;Steidl et al . 2006),这对男性和女性都是相似的(Bradley et al . 2017a)。
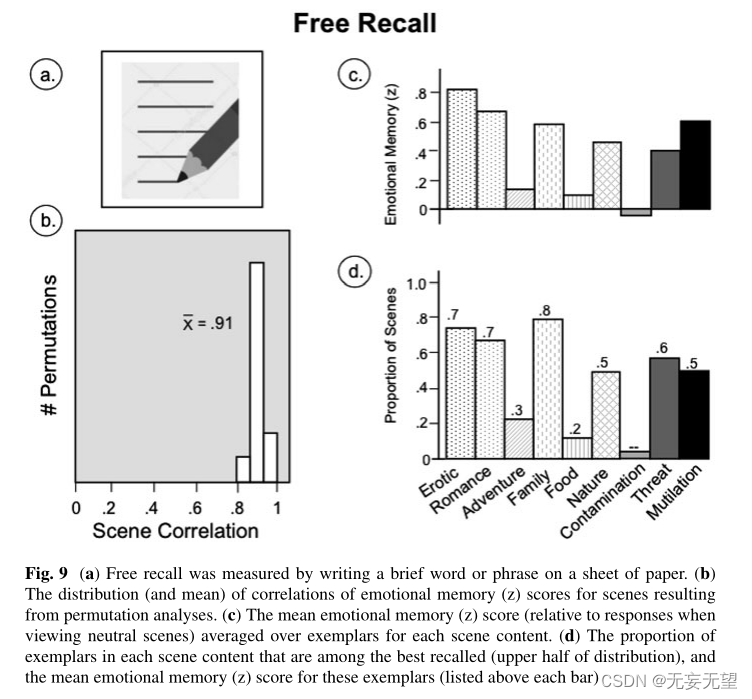
在这里分析的记忆数据中,通过要求每个参与者(在6个不同的组中观看不同的场景中的1个组)写下一个单词或简短的短语(见图9a)来描述每个记住的场景(总共可能有60个),提供足够的信息,使另一个人能够从呈现的场景中识别出范例,从而获得偶然的自由回忆。所有的参与者都能在5分钟的时间内完成自由回忆。
特定图像在自由回忆方面表现出非常高的可靠性,平均场景相关性为0.91 (p < 0.05;见图9b)。图9c显示了特定场景类别的情绪记忆(z)分数(即回忆表现相对于中性场景的回忆),其模式与许多生理测量中获得的模式有所不同。对情色、浪漫、家庭、自然、威胁和残害等场景的记忆表现相当,而对冒险、食物和污染等场景的记忆表现非常差。如图9d所示,在记忆良好的内容类别中,有相当大比例的个体样本被回忆得最好。这些数据与之前的数据一致,这些数据显示,无论是愉快的还是不愉快的场景都能唤起更好的记忆,同时,唤醒程度较低的愉快场景对记忆也有小的好处(Bradley等人2017a),但也表明,特定的场景内容是调节情绪场景自由回忆表现的关键因素。
 6 Discussion
6 Discussion
特定的照片图像是高度可靠的情感反应的激发者在各种3系统的情绪测量参与者。这种力量在很大程度上在于,在随机人群中,引起最大或最小情绪反应的画面是相当稳定的。因此,一个特定的场景往往会引起参与者普遍的情绪反应的增强或减少,而不是对不同个体的特殊吸引力。
此外,对于各种情绪反应,特定场景内容的所有或大部分范例都是最具反应性的,这为基于场景内容选择刺激提供了一些实验确定性。
在情感场景观看期间测量的最可靠(非评估性)数据是皮肤电导和瞳孔直径变化,以及自由回忆。
对于每一项测量,在实验室中,性和暴力场景最一致地引起强烈的情绪反应(以及更好的回忆),其中76人占大多数
在每个测量中,每个内容中最具反应性的范例,与观看场景时相对强烈的食欲和防御动机系统激活相一致。尽管对色情场景(尤其是女性,这可能受到文化或政治规范的影响)的适度唤醒评级,布拉德利等人
2001b),数据表明这些图像强烈而可靠地引起情绪反应。个别科学家或机构审查委员会有时不愿在实验研究中使用或批准这些图像。有趣的是,西海岸(美国)的研究有时倾向于关注呈现暴力图像,而东海岸的研究则更关注露骨的性行为。然而,为了在实验室中有效地研究情绪,必须利用高度激活防御或食欲动机系统的线索,因为在没有这些刺激的情况下,许多依赖的测量将无法显示情绪参与的证据。
在某些测量中,其他场景内容诱发了可靠的情绪反应(参见Bradley等人2001年a,进一步讨论情感场景观看中的内容差异)。例如,皱襞肌肌电(“皱眉”)活动在所有令人不快的场景内容(和大多数范例)中都升高,而只有家庭照片才会引起可靠的颧骨肌电变化。总体而言,心率减速在情绪场景内容中的区别程度低于其他措施,但在唤起愉快和不愉快内容的场景时,心率减速的区别程度最大。杏仁核的激活不仅在色情场景中被提升,而且在浪漫场景中也被提升,浪漫场景描绘了没有明确裸露的爱的互动,这表明在其他生理测量中更仔细地观察了这一内容。
对性爱和浪漫场景的记忆表现也有所提高,但对家庭和自然场景的记忆表现也大幅提高,这使得很难将更好的记忆归因于强烈的情感投入。相反,与先前证明特定图像在记忆表现中的有效性的研究一致,新颖性、独特性、人的存在等其他因素(例如,Bainbridge等2019)可能会影响情绪场景记忆。
综上所述,这些数据确定了适合纳入测量不同情绪反应指数的研究的特定场景内容。
虽然这里没有研究,但在观看场景时,男性和女性在情绪反应上存在一些差异(例如,Bradley等人,2001;芬克等人2017;Sarlo &浮标2017)。男性观看色情作品时的愉悦感和兴奋感通常更高(女性观看暴力作品时更高),而且与女性相比,男性观看色情场景时皮肤电导变化(Bradley等人,2001年b)和瞳孔扩张(Finke等人,2017年)都有所增强。尽管如此,与其他场景内容相比,男性和女性在观看色情场景时都表现出更强的反应,这只是程度上的差异。此外,尽管特定场景是情感投入的可靠诱因,当然并非所有参与者都对所有场景做出反应。虽然一个实验冲动可能是选择倾向于引起强烈情感反应的场景,但另一种观点认为,只包括强烈的情感引发因素可能会减少观察重要个体差异的机会(Lissek等人,2006年),建议包括一个更全面的集合,包括不同的情感反应场景。
图像可以呈现为彩色或灰度,大或小,持续时间短或更长,所有这些都可能影响情绪反应的大小和/或可靠性。先前的研究已经确定,评价等级和情绪反应的生物学测量在图像呈现为彩色还是灰度时变化不大(Bradley等,2001a, 2003;Codispoti等人
2001年,2012年)。此外,在没有视觉掩藏刺激的情况下,短暂的25毫秒呈现继续引发与情感投入一致的各种生理反应(Codispoti et al . 2009)。令人难以置信的是,无论一个场景是全屏呈现还是在矩阵中以多个较小的图像重复呈现以控制局部亮度效果,由情绪唤醒引起的瞳孔后期扩张在幅度上都非常相似(Bradley等人2017b)。这些方法参数的无效效应表明,在成功理解图像的程度上,动机激活继续进行。与这一假设一致,当单个图像的尺寸大大减小(12.5%;codispoi & De Cesarei 2007)或空间模糊(De Cesarei & codispoi 2010),评价评级和指示情感投入的皮肤电反应消失。
图像重复也会对大脑和身体产生实质性的影响,最常见的结果是重复抑制——对重复场景的活动减少。例如,在大脑中,在新场景处理过程中被激活的扩展网络随着场景重复而显着减少,包括杏仁核,尽管情绪和中性场景的差异激活仍然保留了至少4次大规模重复(Bradley et al . 2015)。同样,虽然重复会减少观看场景时皮肤电导的大小和瞳孔直径的变化,但与中性场景相比,情绪场景的差异反应在适度重复后仍然存在(Bradley等人1993;Bradley & Lang 2015)。另一方面,心率显示出相当快的心脏减速下降,并随着重复而完全消失(Bradley等人1993;布拉德利2009)。由于重复对三系统测量的不同影响,在实验研究中选择图像是否重复可能是导致情绪反应大小和稳定性的重要因素。
当前分析中未包括的场景感知过程中情感投入的一种常见测量方法是事件相关电位(ERP;通过放置在头皮上的传感器测量),可以在地形,大小和/或延迟上变化,作为情感场景内容的函数(例如,Schupp et al . 2004)。然而,在单次试验中,erp是出了名的嘈杂,需要大量的试验平均才能获得合理的电压变化估计。而且,由于许多ERP组件对重复特别敏感,因此可以减少延迟和/或振幅(例如,Ferrari等人2011;Codispoti等人2006;法拉利等人(2013),这里进行的单一试验图像分析对于这种情绪处理指数来说不是最佳的。
综上所述,这些数据证实了特定的场景确实会引起个体的情感反应。另一方面,这里进行的初步图像分析可用的数据集缺少一些重要的控制(例如,呈现的场景总数,具体的场景内容,每个内容的样本数量,所有参与者的所有测量的获取)。
未来的研究将解决这些范例差异,并在更大的男性和女性样本(以及整个生命周期)中测量3个系统的反应,这将为科学家提供一个非常有用的数据库,用于选择和平衡最适合特定调查的图像,以及允许更复杂的多变量分析,以确定跨系统情绪反应的具体例子。这样的数据库在开始确定有助于特定示例成功的语义和/或物理特征时也很有用(例如Bainbridge等人2019)。然而,在这样一个生物情感图像集可用之前,本文提供的数据证实了特定场景内容的个体范例可靠地参与了参与者的情感反应,并为选择适合于所调查的情感问题的特定目标和措施的最佳图像集提供了初步的基于数据的信息。
本文来自互联网用户投稿,该文观点仅代表作者本人,不代表本站立场。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。 如若内容造成侵权/违法违规/事实不符,请联系我的编程经验分享网邮箱:veading@qq.com进行投诉反馈,一经查实,立即删除!